要点
- 胃を持たない魚(無胃魚)で共通して欠失・偽遺伝子化した4つの遺伝子を特定。
- 無胃魚で欠失した遺伝子の一部が胃を持たない哺乳類でも欠失していることを確認。
- 器官の喪失に伴う収斂的なゲノム変化の理解や、生物多様性保全への応用に期待。
概要
東京工業大学 生命理工学院 生命理工学系の加藤明准教授、太田地洋大学院生、永嶌鮎美助教、同 科学技術創成研究院 細胞制御工学研究センターの駒田雅之教授、東京大学 大気海洋研究所のSupriya Pipil(スープリヤ・ピピル)博士(研究当時)、渡邊太朗博士、黄國成助教、竹井祥郎名誉教授、静岡大学 学術院理学領域の日下部誠教授、メイヨー医科大学(米国)のMichael F. Romero(マイケル・F・ロメロ)教授らの研究グループは、さまざまな系統に属する無胃魚で共通して4つの遺伝子が欠失・偽遺伝子[用語1]化していることを発見した。
魚類全体の20-27%が胃を持たず、さまざまな系統が独立に胃を失ったことが知られている。胃の喪失に伴う遺伝子欠失として、これまで4つの遺伝子(atp4a、atp4b、pgc、pga2)の欠失が知られていた。今回の研究では、11種の無胃魚(ゼブラフィッシュ、メダカ、ベラ、フグなど)と12種の有胃魚(ナマズ、ニジマス、ティラピア、イトヨなど)のゲノムデータベースを網羅的に比較した結果、新たに4つの遺伝子(slc26a9[用語2]、kcne2[用語3]、cldn18a[用語4]、vsig1[用語5])の欠失・偽遺伝子化を見出すことに成功した。有胃魚(イトヨ)におけるこれらの遺伝子の組織発現を解析したところ、すべて胃に発現していた。さらにこうした遺伝子の一部は、胃のない哺乳類(カモノハシなど)でも欠失していることが確認された。
これらの結果は、器官の喪失に伴ってゲノム中に起きる収斂的[用語6]な変化の理解を前進させるものであり、水生動物の多様な環境適応戦略の理解や生物多様性の保全への応用が期待できる。
本研究成果は、4月3日に「Communications Biology」にオンライン掲載された。

胃を持たない魚類が失った遺伝子を特定
背景
条鰭類[用語7]は44目、453科、約30,000種からなり、魚類最大のグループであるとともに、現存する脊椎動物の半数以上を占める。条鰭類には消化器に胃を持たない系統が存在し、無胃魚と呼ばれる。無胃魚にはコイ目(~3,200種)、ダツ目・カダヤシ目(~1,200種)、フグ目(~3,500種)、ベラ目(~600種)などが含まれる。これらのグループが系統学的に散在していることから、条鰭類はもともと胃を持っていたが、それぞれの系統において胃を喪失したことが分かる。過去の研究では、条鰭類の科の7%、種の20-27%が無胃魚であり、進化の過程で胃の喪失が少なくとも15回にわたって独立に起きたと推定されている(Fish Physiol. 30:1-55, 2010)。胃の喪失はゲノム中の何らかの遺伝子機能の変化により生じるが、胃の喪失自体もさらなるゲノムの変化をもたらすと考えられる。実際に以前の別グループの研究で、さまざまな無胃魚において、胃酸分泌を担うH+ポンプをコードする遺伝子(atp4a、atp4b)や、胃で分泌される消化酵素をコードする遺伝子(pgc、pga2)が欠失していることが報告されている(Proc. Biol. Sci. 281:20132669, 2014)。
これまで東工大とメイヨー医科大学の研究グループはフグ、東大大気海洋研の研究グループはウナギを対象として、魚類の淡水・海水順応に寄与する溶質輸送体26ファミリー(Slc26)の研究をそれぞれ行ってきた。両グループの結果を比べたところ、Slc26の一つであり、ヒトやマウスなどの哺乳類では胃や肺で重要な役割を担うSlc26a9と呼ばれる輸送体について、これをコードする遺伝子(slc26a9)が有胃魚であるウナギ、ニジマス、イトヨには存在し、無胃魚であるフグ、ゼブラフィッシュ、メダカには存在しないことを見出した。ここから、無胃魚で欠失する遺伝子は先に報告されていた4つの遺伝子に限ったものではなく、他にも存在することが示唆されたため、両研究グループは共同で詳細な解析を行った。
研究成果
本研究では、ゲノムデータが公開されている11種の無胃魚(ゼブラフィッシュ、メダカ、ベラ、フグなど)と12種の有胃魚(ナマズ、ニジマス、ティラピア、イトヨなど)を研究対象に選定した。それらの遺伝子構成を比較し、選定した無胃魚の全てもしくは大部分において欠失しているが、有胃魚には存在する遺伝子を網羅的に探索した。その結果、選定した全ての無胃魚において、kcne2、cldn18a、vsig1の3つの遺伝子が欠失・偽遺伝子化していることが分かった。また予備的な解析により数種の無胃魚では存在しないことが既に確認されていたslc26a9は、ベラ目を除く9種の無胃魚で欠失・偽遺伝子化していることが明らかになった(図1)。一方でこれら4つの遺伝子は、選定した全ての有胃魚において無傷な状態で存在していた。さらに、以前の他グループによる研究で欠失が報告されている4つの遺伝子(atp4a、atp4b、pgc、pga2)についても再検証を行ったところ、今回選定した全ての無胃魚において欠失・偽遺伝子化し、全ての有胃魚において無傷な状態で存在していることが示された。これらの結果は、胃の喪失に伴って、slc26a9、kcne2、cldn18a、vsig1、atp4a、atp4b、pgc、pga2の8つの遺伝子の欠失・偽遺伝子化がそれぞれの系統において独立に起きたことを示している。
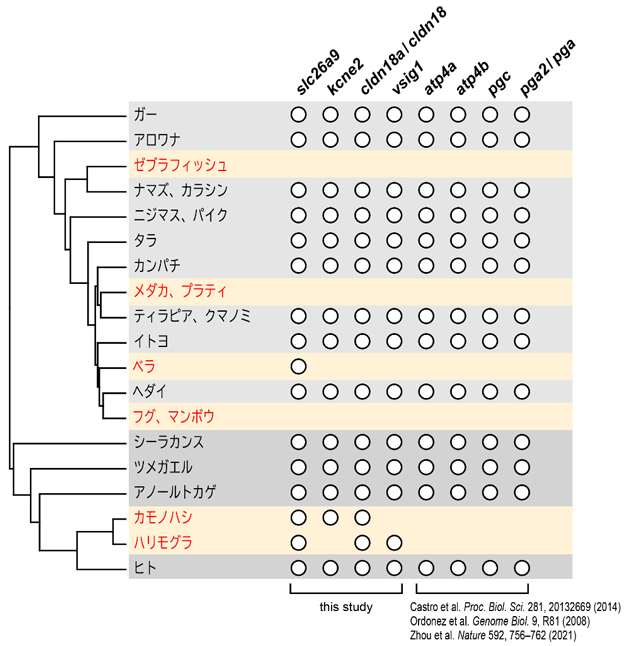
- 図1.
- 無胃魚と単孔類が共通に欠失・偽遺伝子化する遺伝子 胃を持たない、もしくは退化させた系統(無胃魚、単孔類)を赤字で示す。各遺伝子の存在を白丸で示す。本研究ではslc26a9、kcne2、cldn18a(カモノハシ、ハリモグラはcldn18)、vsig1の4遺伝子の有無を解析した。atp4a、atp4b、pgc、pga2(カモノハシ、ハリモグラはpga)の4遺伝子の有無は過去の他のグループによる報告の結果を再検証してまとめた。
今回欠失が確認されたslc26a9、kcne2、cldn18a、vsig1は、哺乳類の胃で重要な機能を担うことが以前から知られているが、魚類における役割は明らかでない。そこで今回、有胃魚のイトヨにおける遺伝子の組織発現をRT-PCR法[用語8]により解析したところ、これら4つの遺伝子の主要な発現器官は胃であることが明らかになった。またin situハイブリダイゼーション法[用語9]によって、イトヨの胃における発現細胞を調べたところ、今回の4遺伝子のうち2遺伝子(slc26a9、kcne2)は以前のatp4a、atp4b、pgc、pga2と同様に主に胃腺細胞に強く発現し、他の2遺伝子(cldn18a、vsig1)は表層上皮細胞に強く発現していた。これらの結果から、無胃魚で欠失している遺伝子が、有胃魚では胃に発現することが確認された。
無胃魚で共通に欠失していた合計8つの遺伝子の機能は、胃酸分泌(slc26a9、kcne2、atp4a、atp4b)、消化酵素(pgc、pga2)、細胞間接着・バリア形成(vsig1、cldn18a)に分類することができる。胃酸分泌に関わる膜輸送体関連遺伝子は他にも存在するが、無胃魚で欠失していた遺伝子は全て、胃の内側の細胞膜で機能するものであった。
原始的な哺乳類である単孔類(カモノハシ、ハリモグラ)も胃を退化させていることが知られる。カモノハシやハリモグラのゲノムデータにおける上記遺伝子の有無を調べたところ、カモノハシではvsig1が偽遺伝子化し、ハリモグラではkcne2が欠失していることが明らかとなった。またカモノハシとハリモグラでは、無胃魚で欠失している8遺伝子のうちatp4a、atp4b、pgc、pga(カモノハシ、ハリモグラにおけるpga2の相同遺伝子)が欠失・偽遺伝子化していることも確認された。
社会的インパクト
器官の喪失という進化上の大きなイベントの仕組みを理解し、生物の生存戦略の違いや消化器の多様性を理解することは、生物多様性のより深い理解や、種の保全、養殖技術の開発などに貢献することが期待される。また、無胃魚で欠失していた遺伝子の一部が胃を退化させた哺乳類でも欠失していたことから、今後これらの遺伝子の解析を通して胃の形成にかかわる未知のメカニズムを解明することで、哺乳類における胃の研究や医学にも貢献できる可能性がある。
今後の展開
本研究では、胃で機能する遺伝子がさまざまな無胃魚の系統において共通して欠失することを明らかにした。しかしながら、転写因子や成長因子の欠失など、胃の喪失の原因として予想される共通のメカニズムを同定することはできなかった。このことは、胃の喪失の原因となるゲノム上の変異が種や系統によって異なる可能性を示唆している。今後は、無胃魚の系統ごとの遺伝子欠失を詳細に解析することにより、胃の喪失のメカニズムを遺伝子レベルでより詳細に明らかにできると期待される。
胃で分泌される酸や消化酵素は、食物を消化するとともに殺菌する作用も持つ。食物の消化は小腸でも行うことが可能であるが、食物を酸で殺菌する役割は他の器官が代わりに行うことはできない。胃の喪失は腸内細菌の獲得に影響を及ぼすことも指摘されており、今後の研究により無胃魚が胃を失ったことによるメリットが明らかになることも期待される。
付記
研究は、日本学術振興会 科学研究費助成事業 挑戦的萌芽研究(24651211)および基盤研究B(26292113、21H02281)の支援を受けて実施された。
用語説明
[用語1] 偽遺伝子 : かつては機能していたが、突然変異や一部配列の欠失により機能を失った遺伝子。さらに時間が経過すると、偽遺伝子の配列全体がゲノムから完全に失われる。これを遺伝子の欠失という。例えばヒトゲノムにはタンパク質をコードする完全な遺伝子が約2万、偽遺伝子が2万以上存在することが知られる。
[用語2] slc26a9 : 溶質輸送体26ファミリー(solute carrier family 26、Slc26)は陰イオン輸送体ファミリーの一つであり、11のメンバーが存在する。slc26a9はその9番目のメンバーであるSlc26a9をコードする遺伝子。Slc26a9は哺乳類の消化管や肺に発現し、Cl-チャネルとして機能する。
[用語3] kcne2 : 電位依存性カリウムチャネルのβサブユニットの一つであるKcne2をコードする遺伝子。αサブユニットの一つであるKcnq1とKcne2は胃の壁細胞(胃酸分泌細胞)の細胞膜上で複合体を形成し、カリウムイオンの分泌に寄与する。
[用語4] cldn18a : クローディン18をコードする遺伝子。クローディンは上皮細胞の密着結合(タイトジャンクション)を形成する膜タンパク質で、上皮組織のバリア形成や細胞間隙を介した物質透過の選択性を担う。クローディン18は水素イオンの細胞間隙の透過を防ぐバリアとして機能する。真骨魚類は全ゲノム重複により生じた2つのクローディン18遺伝子(cldn18a、cldn18b)を有する。
[用語5] vsig1 : 免疫グロブリンスーパーファミリーに属する細胞間接着タンパク質の一つであるVsig1をコードする遺伝子。胃、肺、精巣に発現することが知られる。
[用語6] 収斂(しゅうれん)的 : 異なる系統の生物が、環境要因などにより類似した形質を獲得することを収斂進化と呼ぶ。
[用語7] 条鰭類 : 石灰質の骨を持つ脊椎動物のうち肉鰭類・四肢動物以外の群。魚類の大半を占める。
[用語8] RT-PCR法 : 細胞や組織から抽出したRNAを試験管内で逆転写(RT, reverse transcription)し、その産物を鋳型にしてPCRを行うことで調べたい遺伝子産物を増幅する実験法。細胞や組織において、調べたいRNAが存在するかどうか、すなわちそのRNAをコードする遺伝子の発現がonになっているか、offになっているかを判定することができる。
[用語9] in situハイブリダイゼーション法 : 組織をホルムアルデヒド等で処理し、組織中の様々な細胞に含まれるRNAが分解せず、かつ本来働いている場所から動かない状態に固定する。その後、調べたいRNAと相補的な配列を持つDNAを標識し、細胞内に固定されたRNAと二重らせん構造を形成させて標識することにより、調べたいRNAが組織内で働いている部位を可視化することができる。また、組織内のどの細胞種がそのRNAをコードする遺伝子の発現をonにしているかを特定することができる。
論文情報
掲載誌 : |
Communications Biology |
論文タイトル : |
Convergent gene losses and pseudogenizations in multiple lineages of stomachless fishes |
著者 : |
Akira Kato, Supriya Pipil, Chihiro Ota, Makoto Kusakabe, Taro Watanabe, Ayumi Nagashima, An-Ping Chen, Zinia Islam, Naoko Hayashi, Marty Kwok-Shing Wong, Masayuki Komada, Michael F. Romero, Yoshio Takei |
DOI : |
- プレスリリース 胃を持たない魚類が収斂(しゅうれん)的に失った遺伝子を特定 —器官の喪失に伴うゲノム変化の理解に向けて前進—
- 魚類が「ホウ酸」を体外に排出して 海水に適応するメカニズムを解明|東工大ニュース
- 魚が淡水中のわずかな栄養素を取り込む機構を解明|東工大ニュース
- 加藤明 Akira Kato|研究者検索システム 東京工業大学STARサーチ
- 加藤研究室
- 【研究室紹介】 加藤研究室|生命理工学系 News
- 生命理工学院 生命理工学系
- 細胞制御工学研究センター
- 科学技術創成研究院
- 東京大学 大気海洋研究所
- 静岡大学
- メイヨー医科大学
- 研究成果一覧


お問い合わせ先
東京工業大学 生命理工学院 生命理工学系
准教授 加藤明
Email akirkato@bio.titech.ac.jp
取材申し込み先
東京工業大学 総務部 広報課
Email media@jim.titech.ac.jp
Tel 03-5734-2975 / Fax 03-5734-3661
東京大学 大気海洋研究所 広報戦略室
Email kouhou@aori.u-tokyo.ac.jp
静岡大学 総務部 広報・基金課
Email koho_all@adb.shizuoka.ac.jp
Tel 054-238-5179 / Fax 054-238-4450